Южноафриканский гепард
Acinonyx jubatus jubatus (Schreber, 1775)

Южноафриканский или намибийский гепард.
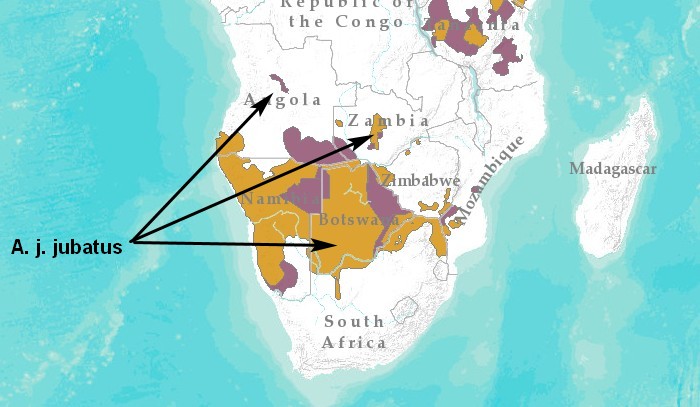
Ареал: Южная Африка (Ангола, Ботсвана, Замбия, Зимбабве, Мозамбик, Намибия и ЮАР). Согласно данным IUCN (2022), окончательно вымершим считается в Малави и Эсватини. Не совсем понятно состояние популяции гепардов на юге Танзании. Сообщалось, что гепарды встречались относительно редко в обширном поясе лесов миомбо на юге Танзании и севере Мозамбика, но в последние годы в этих местах обитания не отмечались. Однако недавнее исследование показало, что леса миомбо могут быть важной средой обитания для гепардов.
Не так давно Kitchener et al. (2017) в подвид включили A. j. raineyi, якобы на основании генетических исследований. Однако более детальное генетическое исследование (Prost et al., 2022) хотя в целом обнаружило близкую связь между гаплотипами обоих подвидов, но доказало их генетическую обособленность. Более ранние исследования предположили, что подвиды разделились, вероятно, во время последнего ледникового максимума (21 тыс. лет назад). Согласно другим авторам, возможно, также в разделении сыграли роль ледники в горах Восточной Африки.
Хотя в современных публикациях (например, Khalatbari et al., 2022, где одним из соавторов является A. C. Kitchener) A. j. raineyi снова признается отдельным подвидом, многие ресурсы, которые опираются на "Википедию" (например, iNaturalist, EOL и др.) продолжают их объединять, что приводит к путанице.

Южная Африка является глобальным оплотом гепарда, где, по предварительным оценкам (IUCN, 2022), насчитывается 3526 взрослых особей, распределенных как минимум по 11 субпопуляциям. Однако вышеуказанная цифра не соответствует приведенной разбивке по субпопуляциям, которая в сумме дает 3650 особей. Крупный центр распространения включает большую часть региональной популяции, около 3396 взрослых особей, распределенных по обширному трансграничному ландшафту, охватывающему южную Ботсвану, Намибию, южную Анголу, северную часть ЮАР и юго-западный Мозамбик. Остальные субпопуляции в регионе намного меньше: 60 половозрелых особей в НП Кафуэ, Замбия; 46 в НП Хванге и его окрестностях, Зимбабве; 42 в НП Гонарежу и заповеднике Сейв, Зимбабве; 37 в трех заповедниках на юге Зимбабве; 18 в Лиува-Плейнс, Замбия; 11 в долине Замбези; 9 в НП Баньине, Мозамбик; 4 в Заповеднике носорогов, Зимбабве; 24 в регионе Мошико в центральной Анголе; и 3 в Матусадоне, Зимбабве. Последняя субпопуляция существенно сократилась после реинтродукции гепарда в середине 1990-х годов и может указывать на плохую долгосрочную жизнеспособность изолированных популяций гепардов на небольших территориях. Большая часть (75%) предполагаемого ареала обитания в регионе находится за пределами охраняемых территорий, на землях, используемых в основном для разведения домашнего скота, а также дичи. Более крупные конкуренты, такие как львы и пятнистые гиены, были истреблены на большей части этого ареала.

Как отмечают сами авторы последнего отчета IUCN (Durant et al., 2022), к представленным оценкам численности гепарда следует относиться с особой осторожностью и они приводятся только в качестве ориентира. Оценки плотности и численности гепарда неточны, и небольшое изменение в оценке средней плотности может привести к значительному общему изменению оценок популяции. Тем более, как указывалось выше, приведение оценки численности 2017 к состоянию 2022 г. с учетом новых данных, на которую опирались авторы проведено не совсем корректно - общая оценка 3526, а сумма перечисленных оценок 11 субпопуляций дает 3650 особей. Разница небольшая, но оценки действительно являются крайне предварительными. Что, впрочем, неоднократно подчеркивают сами авторы.

Самый многочисленный подвид, описание которого обычно приводят как типичное для вида. Общая окраска охристо-желтая иногда темнее или светлее (от коричневато-кремовой до бледно-палевой). Мех обычно немного короче и плотнее, чем у других подвидов. Вдоль плеч и спины обычно заметна небольшая грива. Тело покрыто круглыми черными пятнами, более или менее одинаковыми по размеру, между которыми обычно расположены мелкие пятнышки, из-за чего в целом все пятна кажутся более частыми, чем у большинства других подвидов. Вентральная часть почти белая, с более длинной шерстью и неясными пятнами. От переднего угла глаза до края губы проходит заметная черная линия; ухо с черным пятном сзади и рыжеватым кончиком. Подбородок белый, без пятен; грудь беловатая, пятнистая. Пятна на хвосте к концу образуют 6-8 несовершенных колец к кончику.
Однако южноафриканский гепард занимает территорию с очень разными климатическими условиями. Это, наряду с индивидуальной изменчивостью особей данного вида, приводит к некоторой разнице не только в окраске, но и отличиям в размере и во внешнем виде.
.jpg)
Королевский гепард - знаменитая редкая мутация окраски. Впервые обнаружен в 1926 г. майором А. Л. Купером недалеко от Солсбери на территории современного Зимбабве, поэтому некоторое время назывался по его имени. Был описан как отдельный вид Acinonyx rex (Pocock, 1927) и за гепардом закрепилось новое название - "королевский". Однако несколько позже Покок (1939) признал свою ошибку и согласился, что это цветовая мутация.
Основной цвет преимущественно кремово-охристый; живот белый. По размерам и пропорциям подобен A. j. jubatus, но отличается от него ярким рисунком из черных полос и пятен, продольных на спинной части и косых или продольных на боках; ноги в пятнах до ступней; базальная половина хвоста с двумя продольными полосами, дистальная половина с неправильными поперечными полосами. Длина тела с головой двух описанных Пококом шкур составляла 127-135 см, длина хвоста 76 см.
За исключением одной шкуры, обнаруженной в заповеднике Сингоу в Буркина-Фасо в 1992 г., все шкуры и последующие наблюдения происходят из зоны на востоке и юго-востоке Зимбабве, востоке Ботсване и в провинциях ЮАР Лимпопо и Мпумаланга.

Как указывалось выше, южноафриканский гепард занимает территорию с очень разными климатическими условиями. Это приводит к некоторой разнице не только в окраске, но и отличиям в размере и во внешнем виде. Измерения гепардов в Ботсване (2003-2012) в Южном округе, Кгалагади и Ганзи показали, что свободно перемещающиеся гепарды в соседней Намибии были ниже ростом и стройнее. По сравнению с Намибией, гепарды в Ботсване имели значительно больший обхват груди у самцов и самок, длину хвоста у самцов, общую длину у самцов, а также длину и ширину семенников, но имели значительно меньшую общую длину задних ног у самок.
В Ботсване (Южный округ, Кгалагади и Ганзи) самцы и самки имели следующие размеры, соответственно: длина тела 128,9±7,2 и 122,2±6,2 см; длина хвоста 80,0±3,6 и 73,5±7,2 см; обхват груди 77,1±4,7 и 70,1±3,3 см; общая длина передних ног 77,5±3,7 и 73,3±4,0 см; общая длина задних ног 79,3±4,4 и 74,2±5,5 см; масса 46,7±8,7 и 38,5±3,5 кг. Клыки у самцов были крупнее, чем у самок: длина верхних клыков 2,3±0,против 2 2,1±0,2 см; длина нижних 1,7±0,2 против 1,5±0,2 см.
В Намибии самцы имели в среднем длину тела 125,5 см (108-152 см), длину хвоста 76,7 см (51-87 см), вес 45,6 кг (31-64 кг); самки среднюю длину тела 120 см (105-135 см), длину хвоста 72,5 см (57-79 см), вес 37,2 кг (26-51 кг).
В ЮАР 7 самцов имели длину тела с головой 122,8 см (119–131), длину хвоста 74,4 см (60–84), а вес 4 самцов составлял 55 кг (50–62). В то же время у одной самки был измерен только вес, который составил 57 кг. В НП Калахари Гемсбок самец весил 53,9 кг, а самка 43 кг.

Южноафриканский гепард обитает на лугах, в саваннах, на засушливых участках, в пустынных и полупустынных районах, в скалистых предгорьях. Гепарду необходимы открытые участки, где есть возможность для погони на высокой скорости за травоядными, такими как антилопы. В Южной Африке гепарды встречаются на открытых или слегка поросших деревьями травянистых территориях. В Юго-Западной Африке - на каменистых грядах, грядах песчаного вельда, реже в густой кустарниковой местности, густом сухом лесу, горных пустошах или болотах. На северо-западе ареала раньше отмечался и в лесах миомбо в южной Танзании и северном Мозамбике, однако сейчас там, вероятно, истреблен.
Плотность гепардов (3,2±3,1 против 0,9±0,7 особей/100 км²) более чем в 3 раза выше в Ботсване (Южный округ, Кгалагади и Ганзи), но выборка была небольшая. В другом исследовании (Belbachir et al., 2015) для сельхозугодий Намибии указывается плотность 0,25-0,83 ос./100 км², а в НП Крюгер - 0,88 ос./100 км².

В отличие от Серенгети, размеры участков в лесных саваннах варьируются от 24 км² в Матусадоне в Зимбабве до 320 км² в Калахари. Кочевой характер спрингбоков в Калахари и концентрированное распределение добычи на лугах в НП Матусадоне, вероятно, влияют на размер домашних участков самок гепардов в этих районах. В НП Крюгер, где нет мигрирующей добычи, размеры участков оценивались в 126 км² для коалиции из трех самцов, 195 км² для одиночного самца и 150 км² и 171 км² для двух самок гепарда. Самки чаще использовали более густые леса, чем самцы, поскольку на них, по-видимому, влияло распределение их основной добычи, импал, которые также предпочитают более густые леса.
В Ботсване (Южный округ, Кгалагади и Ганзи) размеры домашних участков более чем в 3 раза меньше, чем в соседней Намибии (517,9±263,4 против 1651,1±1594,2 км²).

Очень большие участки зарегистрированы в Намибии, где также нет мигрирующей добычи. Более раннее исследование указывает 1500 км², что мало отличается от исследования начала 2000-х, которое обнаружило, что в течение жизни у намибийских гепардов средняя площадь участка обитания (метод 95% ядра) составляет 1651 км² (±1594 км²). В отличие от Серенгети, здесь не обнаружено различий между полами или социальными группами по размеру участка, причем все они занимали очень большие территории. У 15 одиночных самцов средний участок составлял 1490 км²; у 11 самцов в коалициях - в среднем 1344 км²; а у 15 самок в среднем 2160 км². Несмотря на такие большие участки, гепарды, как правило, интенсивно использовали лишь небольшую часть этой площади, при этом основные площади (ядра) составляли в среднем 14% от размера всего участка. Участки не были эксклюзивными: самцы демонстрировали большее перекрытие участков между собой, чем самки. В этом случае плотность гепардов и размер их участков в Намибии, по-видимому, определяются не только добычей, но и другими факторами, на которые могут влиять антропогенные воздействия.

Гепарды охотятся, обнаруживая добычу визуально, и в основном днем, особенно с 07:00 до 10:00 и с 16:00 до 19:00, в часы, когда конкуренты менее активны, хотя ночная охота была зарегистрирована в Намибии. Принцип охоты не отличается от таковой у других подвидов - это приближение к добыче (скрытно, открыто или вовсе из засады), преследование на короткой дистанции, сбитие жертвы с ног и удушение (если добыча крупная) или убийство укусом за морду, в заднюю часть шеи, либо проламывание черепа (всё это в случае мелкой добычи).
Добычу либо съедают на месте убийства, либо тащат в ближайшее укрытие. Гепардам обычно требуется время, чтобы прийти в себя после погони, иногда до целого часа. Питаются они быстро, обычно сначала съедая мясо с брюшной поверхности, затем печень и сердце.
В очень редких случаях гепарды едят падаль или возвращаются к своей брошенной добыче, хотя в ЮАР зарегистрированы 12 случаев, когда самки с крупными детенышами возвращались к брошенной добыче после того, как оставили ее на ночь. Имеются предварительные данные о том, что гепарды остаются рядом с убитой крупной добычей, а не бросают ее после насыщения в сельхозугодьях Намибии, где были уничтожены львы и гиены.

Охотится в основном на антилоп среднего размера, от стейнбока (Raphicerus campestris) и дукера до импалы (Aepyceros melampus), спрингбока (Antidorcas marsupialis), редунки (Redunca sp.) и даже куду (Tragelaphus strepsiceros). Когда появляется возможность, убивают овец, коз и страусов. В целом, как и по всему ареалу, гепарды предпочитают убивать наиболее доступную добычу в диапазоне массы тела 23–56 кг с пиком в 36 кг. Как и везде, предпочтение отдается молодым животным, а не взрослым особям, особенно в случае более крупных видов, таких как обыкновенные антилопы гну (Connochaetes taurinus).
В целом, на юге Африки основной добычей являются спрингбоки (северо-восток Ботсваны; НП Калахари Гемсбок, ЮАР; НП Этоша, Намибия); телята куду и бородавочники (Phacochoerus africanus) (ранчо Намибии). Важной добычей являются орикс (Oryx gasella) и беломордый бубал (Damaliscus pygargus).
В лесных массивах саванны НП Крюгер импалы являются доминирующей добычей (68% из более чем 2500 убитых), хотя гепарды в этом регионе отдают предпочтение редунке (Redunca arundinum). В охотничьем заказнике (ОЗ) Пхинда в провинции Квазулу-Натал, где гепарды были повторно интродуцированы, наиболее частым видом добычи была ньяла (Tragelaphus angasii), самое многочисленное доступное копытное среднего размера, на которое приходилось 39% всех добытых животных, за ней следовала импала (34,5%). В Калахари преобладающей добычей являются спрингбоки (86,9% добычи), тогда как в НП Кафуэ в Замбии чаще всего добывались пуку (Kobus vardonii).

Обычно гепарды выбирают копытных, которые находятся поодиночке или небольшими группами, в высокой растительности, менее бдительны, на краю стада и вдали от ближайших соседей. Самцы ловят более крупную добычу, чем самки, из-за их большего размера и охоты коалициями. В ОЗ Пхинда более 20% добычи, убитой парами и тройками самцов, весили более 100 кг, тогда как только одна добыча (менее 1% от общего количества), добытая самками, весила более 100 кг. Здесь же наблюдалось, как коалиции самцов дважды захватывали молодых жирафов (Giraffa camelopardalis). Однако взрослые самцы не обязательно более успешны при охоте группами, чем поодиночке. Гепарды иногда получают травмы во время охоты, хотя смертельные случаются редко.
Следует отметить, что чаще всего самцы из коалиции редко помогают друг другу при захвате добычи; чаще один просто начинает охоту, а остальные следуют за ним. Лишь крайне редко они могут преследовать добычу совместно, но и тогда эта охота не напоминает коллективную, которая вообще не свойственна кошачьим, даже львицам из одного прайда. Точно такая же ситуация как с самцами, происходит и с самками, которые обучают охоте своё еще не отделившееся полувзрослое потомство. Чаще молодые не помогают матери при охоте, а мешают.

В НП Ливонде, Малави, в рационе реинтродуцированных гепардов зарегистрировано 13 видов добычи. Гепард отдавал наибольшее предпочтение большому куду (Tragelaphus strepsiceros) при рассмотрении популяций добычи. Четыре вида составляли основную часть рациона гепардов, а именно; куду, импала (Aepyceros melampus), водяной козел (Kobus ellipsiprymnus) и бушбок (Tragelaphus sylvaticus). Кроме того были зарегистрированы кустарниковый дукер (Sylvicapra grimmia), канна (Taurotragus oryx), черная антилопа (Hippotragus niger), стенбок Шарпа (Raphicerus sharpei), большой редунка (Redunca arundinum) и суни (Neotragus moschatus). А также кустарниковая свинья (Potamochoerus larvatus), кустарниковый заяц (Lepus saxatilis) и неопределенный представитель пресмыкающихся из отряда Squamata.

Хотя основная добыча гепардов копытные, естественно, что хищник добывает не только самую многочисленную и оптимальную добычу, но и факультативно всех тех, которые доступны в данное мгновение. Это касается павианов, наземных птиц (преимущественно крупных), зайцев и дикобразов и многочисленных грызунов. Кроме того, в некоторых регионах нетипичная добыча может стать важной для местных гепардов, например, в некоторых частях Намибии гепарды полагаются на зайцев и других грызунов, когда альтернативная добыча недоступна.
Гепарды также могут убивать и есть других хищников, таких как шакалы (Canis spp. и виды Lupulella) и лисиц (Vulpes spp.). Имеется одна необычная запись о том, как самка гепарда пыталась убить молодого медоеда (Mellivora capensis) в частном охотничьем заказнике Нгала в провинции Лимпопо, Южная Африка. Драка закончилась, когда на помощь пришла вторая особь медоеда, возможно, мать первого.

В Намибии гепард является важным хищником, истребляющим домашний скот как на коммерческих, так и на общинных фермах. Хищничество гепардов в сельскохозяйственных угодьях Намибии составляет не менее 0,01 теленка и 0,004 овцы на км² или в среднем 0,76 телят и 0,32 овцы в год. Однако в цифрах ежегодные потери на этих фермах составляют 10-15% для мелкого поголовья (овцы и козы) и 3-5% для телят крупного рогатого скота до 8-месячного возраста.
Гепарды хорошо адаптированы к жизни в окружающей среде. Они могут долго обходиться без воды и, по оценкам, в пустыне Калахари проходят в среднем 82 км между водопоями. Замечено, что гепарды удовлетворяют свои потребности во влаге, выпивая кровь или мочу своей добычи или поедая дыни тсамма (дикий южноафриканский арбуз).
Гепарды практически не пытаются защитить свою добычу от более крупных хищников и клептопаразитов, таких как львы (Panthera leo) и пятнистые гиены (Crocuta crocuta). Как следствие, гепарды избегают охоты в районах присутствия этих хищников. Зарегистрированы случаи, когда стервятники леопарды (Panthera pardus), гиеновидные собаки (Lycaon pictus), бурые гиены (Hyaena brunnea) и даже павианы (Papio ursinus) отгоняли гепардов от добычи.

Социальная структура обычная для всех гепардов, т. е. самки либо одиночные, либо с потомством, а самцы также либо одиночные, либо создают коалиции из 2-3 (реже 4) особей. Большинство этих коалиций создаются на всю жизнь и полностью состоят из однопометников, но нередко могут включать неродственного самца. Обычно коалиция территориальна, т. е. занимает определенный участок, который защищается от других самцов. У одиночных самцов система сложнее. Они могут быть территориальными (резидентами) или бродячими. Причем, это может длиться как всю жизнь, так и меняться с течением времени. Зафиксировано много случаев, когда наблюдаемые долгое время гепарды как самцы-резиденты становились бродячими, и наоборот, отдельные особи, побродив (иногда длительное время, иногда недолго), обзаводились личными домашними участками.

Что касается самок, то достигнув репродуктивного возраста, они практически все становятся территориальными, причем многим свойственна филопатрия - создание собственного участка либо рядом с натальным участком (материнским), либо даже непосредственно на нем. Однако территориальность самок довольно условная, их участки широко пересекаются с участками других самок и самки просто избегают друг друга по времени посещения определенного района. Это же касается и самцов или их коалиций, которые поддерживают непосредственную близость к одиночной или самке с детенышами, когда у той начинается течка. Такая ассоциация может длиться до 2 дней, хотя имеются наблюдения, когда самцы общались с самками без эструса и их детенышами в течение до 18 дней. Такие ассоциации могут объяснять наблюдения крупных по размерам групп гепардов. Большие группы до 14-19 животных (включая детенышей) время от времени регистрировались в некоторых частях южной Африки, где были истреблены другие крупные хищники (Ботсвана, Намибия). Преимущества группировки в таких условиях не ясны.

Размер помета и остальные параметры обычно зависят от условий проживания гепардов. Например, имеются сравнения данных из различных заповедников ЮАР (2011), где в 4 присутствовали львы, а в 3 отсутствовали. Размер помета при выходе из логова у гепардов составлял 3,8 против 4,6; число доживших до независимости молодых из помета - 2,9 против 4,7; возраст достижения независимости 18,8 против 14,9 мес. На охраняемых территориях при наличии львов, леопардов и др. хищников в НП Пиланесберг (2019) и заповеднике Мун-Я-Вана (2020), в который входит ОЗ Пхинда, возраст достижения независимости составлял 16,2 и 16,7 мес соответственно.
Уровень смертности котят до обретения независимости может быть высоким, отчасти из-за высокой смертности в период от рождения до 3-месячного возраста. В 1960-х сообщилось о 50% смертности котят в НП Крюгера, а в 1990-х о 38% смертности в ОЗ Пхинда. В Трансграничном парке Кгалагади выживаемость детенышей от рождения до обретения независимости составила 35,7%. В НП Пиланесберг (2019), где обитает много львов и леопардов, смертность котят гепарда составляет ок. 50%. При гибели помета самки быстро возобновляют течку и снова беременеют (в среднем, через 19 дней). Детоубийство, т. е. убийство детенышей чужими самками или самцами гепардов у этого вида не отмечено.

По данным из Намибии, самцы и самки достигают взрослой массы тела после 49-месячного возраста. В Ботсване в возрасте более 12 месяцев самцы, как правило, крупнее самок по массе тела, длине тела и обхвату груди, но не по общей длине передних ног. Самки достигают своего полного размера в возрасте 31–48 месяцев, тогда как самцы - только в возрасте старше 48 месяцев.
Молодые могут не отделяться от матери, пока им не исполнится 15–17 месяцев. Затем они образуют группы, но самки обычно покидают их перед наступлением первой течки (в 17-27 месяцев). Возраст первого размножения у самок в заповедниках ЮАР в среднем составляет 27-28 мес.
В начале самостоятельности способности молодых гепардов преследовать добычу на большой скорости и сбивать ее с ног развиты слабо, поэтому они часто вынуждены довольствоваться более мелкой добычей, а в некоторых местах первое время живут впроголодь. В таких условиях молодым самкам сложно достичь кондиций, оптимальных для размножения и выращивания потомства. Поэтому чем больше конкурентов, а значит и доступной добычи, тем позднее самки приступают к размножению. Хотя всё это индивидуально и часто разница в возрасте либо незначительная, либо совсем обратная. Так, например в заповедниках ЮАР (2011), где отсутствуют львы, возраст первого размножения самок составил в среднем 28,2 мес, а где были львы - 26,9 мес.

Южноафриканский гепард является уязвимым подвидом в следствии браконьерства, охоты, потери среды обитания и отсутствия добычи. Хотя хищничество против детенышей гепарда может быть чрезвычайно высоким, взрослые гепарды относительно невосприимчивы к хищникам, хотя сообщалось о редких случаях нападения львов, гиен и леопардов. Львы, пятнистые гиены и леопарды являются основными убийцами детенышей гепарда, хотя свою роль играют и более мелкие хищники, такие как медоеды, шакалы и птицы-секретари (Sagittarius serpentarius).
Систематических исследований болезней диких популяций гепардов не проводится. В Намибии у них практически нет заболеваний, хотя из 7 гепардов НП Этоша, зарегистрированных телеметрическим методом, 6 умерли от сибирской язвы. Однако низкая плотность гепарда делает маловероятным то, что инфекционные заболевания представляют серьезную угрозу для свободно перемещающихся популяций гепардов. Вирус иммунодефицита кошек в Набибии практически отсутствует (0–2%).

В Зимбабве ареал гепардов сокращался на 11% в год в период с 2007 по 2015 год и ныне этот вид исчез из 63% своего ареала за последние 10–15 лет. Популяция сократилась на 85% за аналогичный период: с предполагаемого минимума в 1520 гепардов в 1999 г. до всего лишь 150–170 гепардов в 2015 г. Почти все эти потери произошли за пределами охраняемых территорий.
Более того, гепарды истреблены на большей части территории Мозамбика, Замбии, Замбии и Анголы, скорее всего, всего за последние несколько десятилетий. Популяция гепардов во многих странах сталкивается с резким сокращением из-за быстрого изменения землепользования и растущей нетерпимости к гепардам за пределами охраняемых территорий, а также массовой торговли мясом диких животных, которая приводит к исчезновению добычи на больших территориях ареала гепарда.

Как и везде, основным врагом гепарда остается человек, в меньшей степени напрямую, но в огромной - косвенно. Вторжение в среду обитания и сокращение численности копытных, вызванное выращиванием сельхозкультур, являются причиной большей части массового сокращения ареала. Прямое преследование по-прежнему происходит в большинстве районов, где гепарды обитают за пределами охраняемых территорий, например, в Намибии, где подавляющее большинство животных (более 90%) обитает на сельскохозяйственных угодьях. 79,4% зарегистрированных случаев смерти диких гепардов связано с человеком, большинство из которых приходилось на взрослых взрослых животных в возрасте около 5–6 лет. Тем не менее, в период с 1991 по 1999 год ежегодные подсчеты гепардов, убитых фермерами, значительно снизились. Хотя гепарды на сельскохозяйственных угодьях Намибии по-прежнему воспринимаются как проблема, терпимость фермеров к ним, возможно, улучшилась.
Высокоскоростные дороги также представляют растущую угрозу для популяций гепардов. Это вызывает особую озабоченность там, где асфальтированные дороги пересекают или примыкают к основным территориям охраняемой дикой природы. Сообщалось также о смертях на многих других дорогах, в том числе в ЮАР и Замбии.

Гепарды плохо разводятся в неволе. Южноафриканские гепарды являются наиболее распространенным подвидом для разведения в неволе по всему миру - в Америке, Африке, Евразии и Австралии. Смертность взрослых в неволе составляет 66% смертности и происходит главным образом из-за почечной недостаточности и заболеваний печени. Смертность среди молодых обусловлена различными причинами, но в первую очередь пневмонией.
Размножение в неволе ограничено относительно небольшим количеством эффективных особей (18,2%). Причины неудач разведения в неволе включают инбридинг, неадекватную социальную среду, возможные физиологические проблемы и болезни.
Искусственное оплодотворение оказалось относительно успешным, и самки, зачатые с его помощью, также успешно размножались. Для воспроизводства особенно использовалась сперма намибийских гепардов. Однако спорадические попытки реинтродукции особей, рожденных в неволе, которые предпринимались в районах Южной Африки, в основном потерпели неудачу из-за неспособности этих гепардов охотиться на диких копытных и нападения львов; напротив, перемещение гепардов из дикой природы в дикую природу в ЮАР и Намибии привело к успешному восстановлению отдельных субпопуляций.

В Малави в целях реинтродукции в НП Ливонде с июня 2017 по февраль 2018 были выпущены 7 гепардов (4 самца и 3 самки), которые отслеживались по июль 2019 г.
Для переселения использовались гепарды, отловленные в 5 разных заповедниках ЮАР, причем 3 гепарда были однопометники в возрасте 22 мес (самка CF3 и ее братья CM3 и CM4). Все животные были половозрелые, 6 молодые (22-26 мес) и один самец (CM2) постарше - 78 мес.
До выпуска животные содержались во временных вольерах (бома, 50 х 50 м) разный период времени (23-58 дней), который зависил от прекращения демонстрации стрессового поведения. Затем их выпускали, а ворота бомы закрывали. После выпуска гепардов не кормили. Все самки и 2 самца (CM1 и CM2) были оснащены спутниковыми ошейниками GPS, а один самец в коалиции двух родственных самцов (CM3) был оснащен ошейником слежения УКВ.
К сожалению, судьба гепардов-однопометников сложилась крайне неудачно. Самка CF3 погибла в старой проволочной петле для копытных через 307 дней после выпуска, все 3 ее котенка умерли после смерти матери. Ее братья самцы CM3 и CM4 были зарегистрированы как отсутствующие через 85 и 152 дня после выпуска соответственно.

Все 5 гепардов с GPS-ошейниками в НП Ливонде обустроили свои домашние участки в течение разного времени. Самки обосновались в течение 29-73 дней после выпуска, причем CF3 раньше всех, а CF1 последняя. Во время поисков они посещали значительно большие территории: CF1 до 60 км², остальные около 40 км², однако после обоснования посещаемые участки сократились до 10 км² и менее.
Самцы заняли постоянные участки лишь через 174 дня, после первых обширных исследовательских перемещений. Во время поисков посещаемая территория у самцов CM1 и CM2 достигала 400 и 300 км², соответственно, а после обоснования постоянных участков сократилась до 100 и 50 км². Здесь следует отметить, что самец CM2 был самый старый из всех переселенных особей (6,5 лет).
Однако измерить окончательные домашние участки удалось только для CM1 и трех самок. Домашние участки варьировались от 41 до 97 км², а основная площадь использования - от 13 до 28 км². Все домашние участки и их основные области пересекались. CF3 и CF2 имели самый высокий процент перекрытия участков (71,1%), тогда как CF2 и CF1 имеют самую высокую площадь перекрытия (43 км2). Участок CM2 перекрывался с участками всех самок на 40,1-60,9 %. В основных частях участков наблюдалось одинаковое перекрытие, при этом CM2 и CF2 имели наибольшее перекрытие - 62,7% (16 км²).
Когда самец CM2 постарел и CM1 достиг зрелости, более молодой самец начал вторгаться на территорию старого и в конечном итоге завладел ею после смерти CM2.

Все три самки родили свой первый помет после создания домашнего участка в среднем через 131 день после выпуска (диапазон 123–138 дней). За 2-летний период наблюдения зарегистрировано четыре события родов. Логова располагались в пределах 2,2±0,5 км от бомы. Выращивание в логове котят всех 4 пометов длилось примерно одинаково (53-56 дней). Средний размер помета составлял 4 детеныша (от 3 до 6), что соответствовало средним размерам помета, зарегистрированным в исходных популяциях (CF1 - 3 котенка, CF2 - 4 и 6 котят, CF3 - 3 котенка). Считается, что первые три помета в парке произошли от старого самца CM2.
Все котята достигли возраста выхода из логова, но смертность детенышей была самой высокой в течение первых 6 месяцев, при этом общая выживаемость детенышей составила 60%. Из четырех пометов один полностью погиб после смерти матери (CF3), 6 молодых из двух пометов достигли независимости, а один помет с 6 потомками (младше 12 мес) все еще находился на иждивении на момент исследования. В конце двухлетнего периода наблюдений в 2019 г. популяция состояла из 1 взрослого самца, 2 взрослых самок, 4 полувзрослых самцов, 2 полувзрослых самок и 6 зависимых детенышей.
Насколько успешной будет программа реинтродукции пока судить сложно. Однако на данный момент на сервисе iNaturalist очень много фото гепардов из НП Ливонде. Причем, большинство наблюдений датированы 2021-2022, а некоторые 2023 г., например, фото внизу молодой самки, которая указана как CH39. Следует отметить, что родившиеся в Ливонде гепарды изначально обозначались Ch#.

Международная торговля мехом, преобладавшая в 1960-х годах, на юге Африки практически прекратилась. С 1986 г. вид был классифицирован IUCN как уязвимый, однако это не касалось популяций Ботсваны, Зимбабве и Намибии. Охота запрещена: Ангола, Ботсвана, Малави, Мозамбик, ЮАР. На заседании CITES в 1992 г. была принята годовая квота, разрешающая добывать 150 живых животных или охотничьих трофеев в Намибии, 50 в Зимбабве и 5 в Ботсване. Однако квота - это максимум.
С 2002 по 2011 г. легальная торговля дикими гепардами в среднем составляла 153 экземпляра в год (в основном охотничьи трофеи из Намибии) и 88 живых животных, выращенных в неволе (в основном из ЮАР).
Намибия, ЮАР и Зимбабве с 1990-х гг. реализуют стратегию разрешения трофейной охоты на гепардов на частных землях с целью побудить землевладельцев принимать обитание гепардов на своей земле и получать от них прибыль. Кроме того, Фонд охраны гепардов Намибии работает над обучением фермеров соответствующим мерам управления, которые можно предпринять, чтобы минимизировать потери поголовья.

Литература
1. Krausman P. R., Morales S. M. "Acinonyx jubatus" / Mammalian Species. No. 771, pp. 1-6. Published 15 July 2005 by The American Society of Mammalogists.
2. Pocock R. I. "Description of a new species of cheetah (Acinonyx)". Proceedings of the Zoological Society of London, 1927, 97 (1): 245-252.
3. Pocock R. I. "Acinonyx jubatus (Schreber). The Hunting Leopard or Cheetah" / The fauna of British India, including Ceylon and Burma. Mammalia. Volume 1. London: Taylor and Francis, 1939. pp. 324-325.
4. Harper F. "South African Cheetah (Acinonyx jubatus jubatus)" / Extinct and vanishing mammals of the Old World. Special publication No. 12, New York, 1945. p. 280.
5. Harper F. "King Cheetah; Cooper's Cheetah (Acinonyx jubatus rex)" / Extinct and vanishing mammals of the Old World. Special publication No. 12, New York, 1945. pp. 286-288.
6. Durant S. M., Groom R., Ipavec A., Mitchell N., Khalatbari L. "Acinonyx jubatus" / The IUCN Red List of Threatened Species 2022: e.T219A124366642.
7. Kitchener A. C., Breitenmoser-Würsten Ch., Eizirik E., Gentry A., Werdelin L., Wilting A., Yamaguchi N., Abramov A. V., Christiansen P., Driscoll C., Duckworth J. W., Johnson W., Luo S.-J., Meijaard E., O’Donoghue P., Sanderson J., Seymour K., Bruford M., Groves C., Hoffmann M., Nowell K., Timmons Z., Tobe S. "A revised taxonomy of the Felidae. The final report of the Cat Classification Task Force of the IUCN/SSC Cat Specialist Group" / Cat News Special Issue 11, 2017. - 80 pp.
8. Prost S., Machado A. P., Zumbroich J., Preier L., Mahtani-Williams S., Guschanski K., Brealey J. C., Fernandes C., Vercammen P., Godsall-Bottriell L., Bottriell P., Dalton D. L., Kotze A., Burger P. A. "Conservation Genomic Analyses of African and Asiatic Cheetahs (Acinonyx jubatus) Across Their Current and Historic Species Range" / Molecular Ecology, 2022 Aug; 31(16):4208-4223.
9. Khalatbari L., Durant S., Kitchener A. C., Breitenmoser-Würsten C., Ostrowski S., Brouwer E., Breitenmoser U. "The situation of the Cheetah in Asia". CMS, Bonn, Germany, 2022. - 136 pp.
10. Caro T. "Acinonyx jubatus. Cheetah" / in Kingdon J. & Hoffmann M. (eds) "Mammals of Africa. Volume V. Carnivores, Pangolins, Equids and Rhinoceroses". Bloomsbury Publishing, London, 2013. pp. 187-196.
11. Nowel K., Jackson P. "Cheetah, Acinonyx jubatus, Schreber, 1776" / Wild Cats: Status Surway and Conservation Action Plan. IUCN/SSC Cat Specialist Group. Gland, Switzerland, 1996. pp. 12-16.
12. Boast L. K., Houser A. M., Good K., Gusset M. "Regional variation in body size of the cheetah (Acinonyx jubatus)" / Journal of Mammalogy, 94(6): 1293–1297, 2013.
13. Belbachir F., Pettorelli N., Wacher T., Belbachir-Bazi A., Durant S. M. "Monitoring Rarity: The Critically Endangered Saharan Cheetah as a Flagship Species for a Threatened Ecosystem" / PLoS One. 2015; 10(1): e0115136.
14. Castelló J. R. "Felids and Hyenas of the World: Wildcats, Panthers, Lynx, Pumas, Ocelots, Caracals, and Relatives" Princeton University Press, 2020.
15. Sievert O., Fatterbert J., Marnewick K., Leslie A. J. "Assessing the success of the first cheetah reintroduction in Malawi" / Oryx, 56(4): 1-9, 2022.
16. Bissett C., Bernard R. T. F. "Demography of cheetahs infenced reserves in South Africa: implications for conservation" / South African Journal of Wildlife Research, 2011, 41: 181-191.
17. Sievert O. "Early Post-Release Movements, Prey Preference and Habitat Selection of Reintroduced Cheetah (Acinonyx jubatus) in Liwonde National Park, Malawi" / Thesis presented in partial fulfilment of the requirements for the degree of Master of Science at Stellenbosch University. 2020. - 132 pp.